
The Arctic Cognition Gene, 13 Years Later
Cold winters, seasonality, farming, and the strange history of rs4680

In 2013 I published a paper about a small genetic variant with a large theoretical shadow: COMT Val158Met, also known as rs4680 (Piffer, 2013). At the time, what caught my attention was a peculiar geographic pattern. The Met(A) allele reached some of its highest frequencies in northern Europeans, but also among Inuit populations.
That was an unexpected combination. Northern Europeans and Inuit are not especially close genetically, linguistically, or culturally. What they do share is a harsh ecological reality: long winters, strong seasonality, and environments in which planning failures can be costly.
This observation led me to a hypothesis. Cold, seasonal environments should reward planning. They make food availability less continuous. They raise the value of storage, reliable tools, delayed returns, long-range coordination, and technologies that cannot afford to fail. If the Met(A) allele pushes cognition toward greater stability, working memory, and maintenance of task goals, then colder and more seasonal ecologies might have favored it.
There was also a second hypothesis. Farming is a delayed-return system. It requires storing seed, planning across seasons, defending land, coordinating labor, and accepting more formal rules than mobile foraging usually requires. If the same cognitive style helped with farming, then farmers should have higher Met(A) frequencies than hunter-gatherers.
COMT matters because it helps regulate catecholamines, including dopamine, and it is especially relevant in the prefrontal cortex, where dopamine transporters are relatively sparse. The Val158Met variant is functional: Val is associated with higher COMT activity, while Met(A) produces a lower-activity enzyme (Lachman et al. 1996; Chen et al. 2004). The usual cognitive story is that Met(A) can favor working memory and stable cognitive control, while Val(G) can favor flexibility under some conditions (Egan et al. 2001). But the cognitive literature is mixed, and meta-analytic work cautions against treating rs4680 as a simple intelligence switch (Barnett et al. 2008).
At the time, the available evidence was thin. There were no large ancient-DNA panels to lean on. The argument had to be made from modern population frequencies, ecological proxies, and a theoretical model about climate, cognition, and culture.
Thirteen years later, we can revisit those ideas with thousands of ancient genomes, paleoclimate reconstructions, and genome-wide ancestry controls.
The results are not what I expected.
What Changed
I used AADR v66 2M and extracted rs4680, treating the A allele as the Met allele. For ancient samples, I merged the allele counts with AADR metadata and with CHELSA/TraCE21k climate reconstructions. The main climate variables are coldest-quarter temperature, converted where needed into coldness, and quarter temperature seasonality, the contrast between the warmest and coldest quarters.
For global ancient climate models I used projected genome-wide PCs, PC1 through PC10, plus sample date and 2M coverage. For modern samples I re-geocoded obvious diaspora labels where possible, and also ran a strict version excluding admixed populations without a single homeland coordinate.
The Hypotheses
Table 1. The old hypotheses and the updated ancient-DNA result.
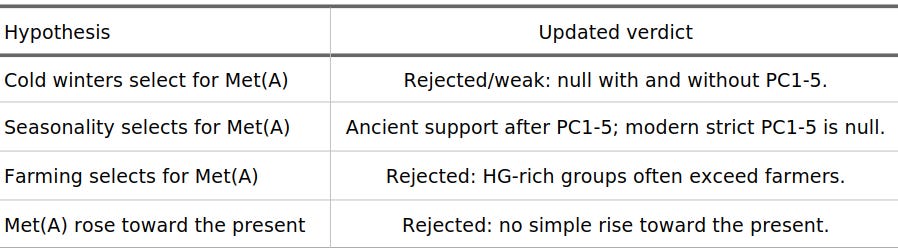
Highest Ancient Groups
Before collapsing samples into hunter-gatherer, farmer, and steppe categories, it is useful to look at the highest-frequency ancient labels directly. The plot below uses N >= 15. Small labels are pooled only when they share a clear geographic/cultural label; here that adds an Old Bering Sea / Chukotka bin.
Figure 1. Highest ancient Met(A) frequencies. Points are sized by sample count; the pooled Arctic/Beringian bin is marked separately.
Two of the most conspicuous high-frequency labels need a little interpretation. Serbia Iron Gates is not a farmer signal: it is a Danube Gorge hunter-gatherer context with eastern hunter-gatherer and steppe-adjacent affinities, sitting on the corridor between the Balkans and the Pontic-Caspian world. Russia_Tatarstan_Eneolithic is also not Yamnaya proper. It is a Volga-Kama Eneolithic group from Murzikha-2, chronologically earlier than Yamnaya but geographically and genetically close to the northern forest-steppe/EHG sphere. In both cases, the high Met(A) frequency points back toward forager and steppe-adjacent population history, not toward the arrival of farming. Curiously, Roman IA/Republican samples also have quite high A frequency.
Hunter-Gatherers Versus Farmers
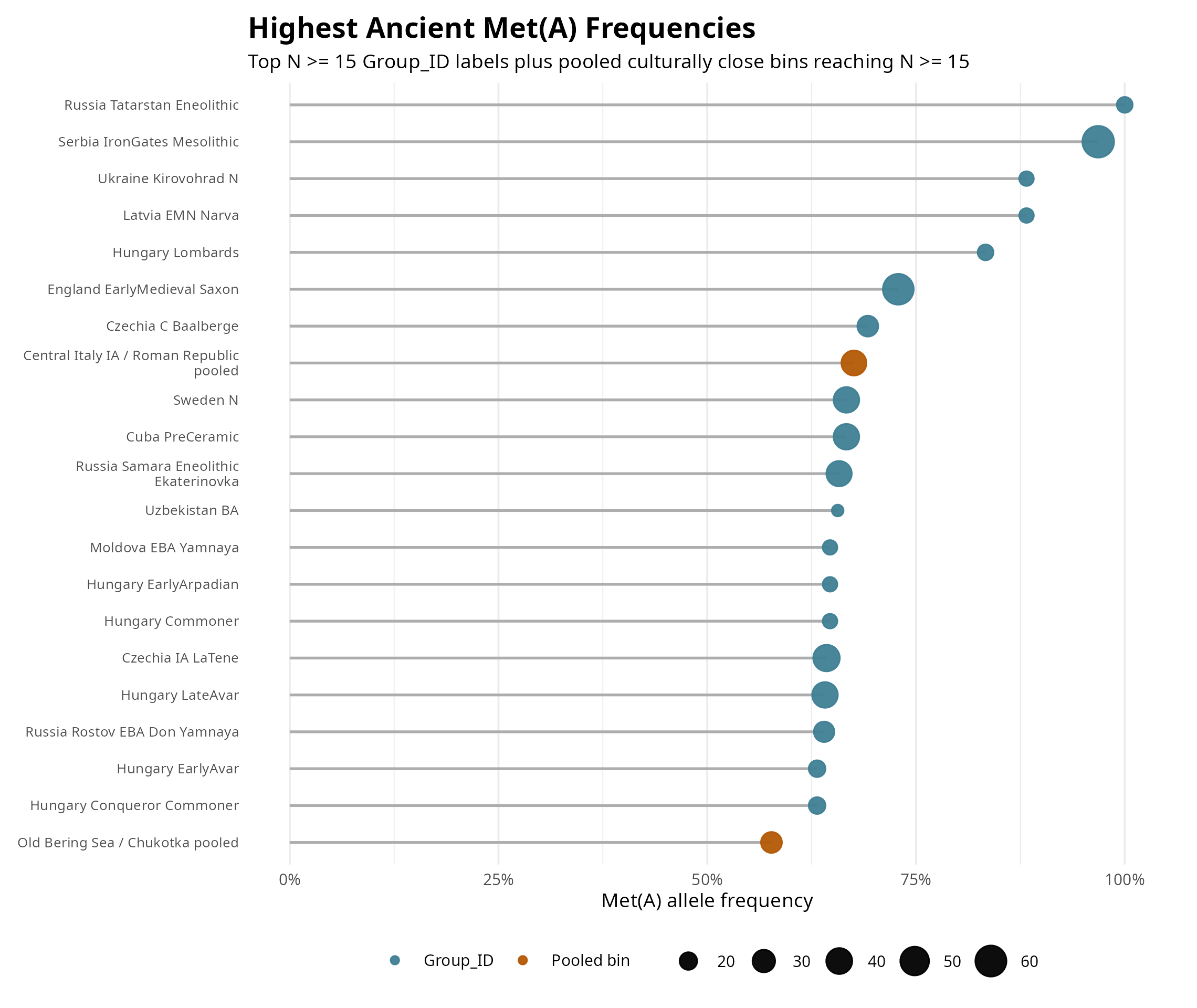
The first result is the most damaging to the farming hypothesis. Ancient hunter-gatherer-rich groups are not low in Met(A). Several are very high. Iron Gates hunter-gatherers, Baltic hunter-gatherers, core WHG groups, and EHG or steppe-forager-like groups all sit above early Anatolian and European farmer groups. Yamnaya is elevated, but not uniquely so. The signal is older and broader than Yamnaya.
That reverses the simple modern farmer-vs-hunter-gatherer contrast from the earlier paper. With ancient DNA, Met(A) does not look like a farming allele. It looks like an allele that was already common in parts of the northern and western Eurasian forager world, then carried forward through later mixtures (Fig. 2).
Figure 2. Met(A) frequency in hunter-gatherer-rich, steppe, and farmer-related ancient groups.
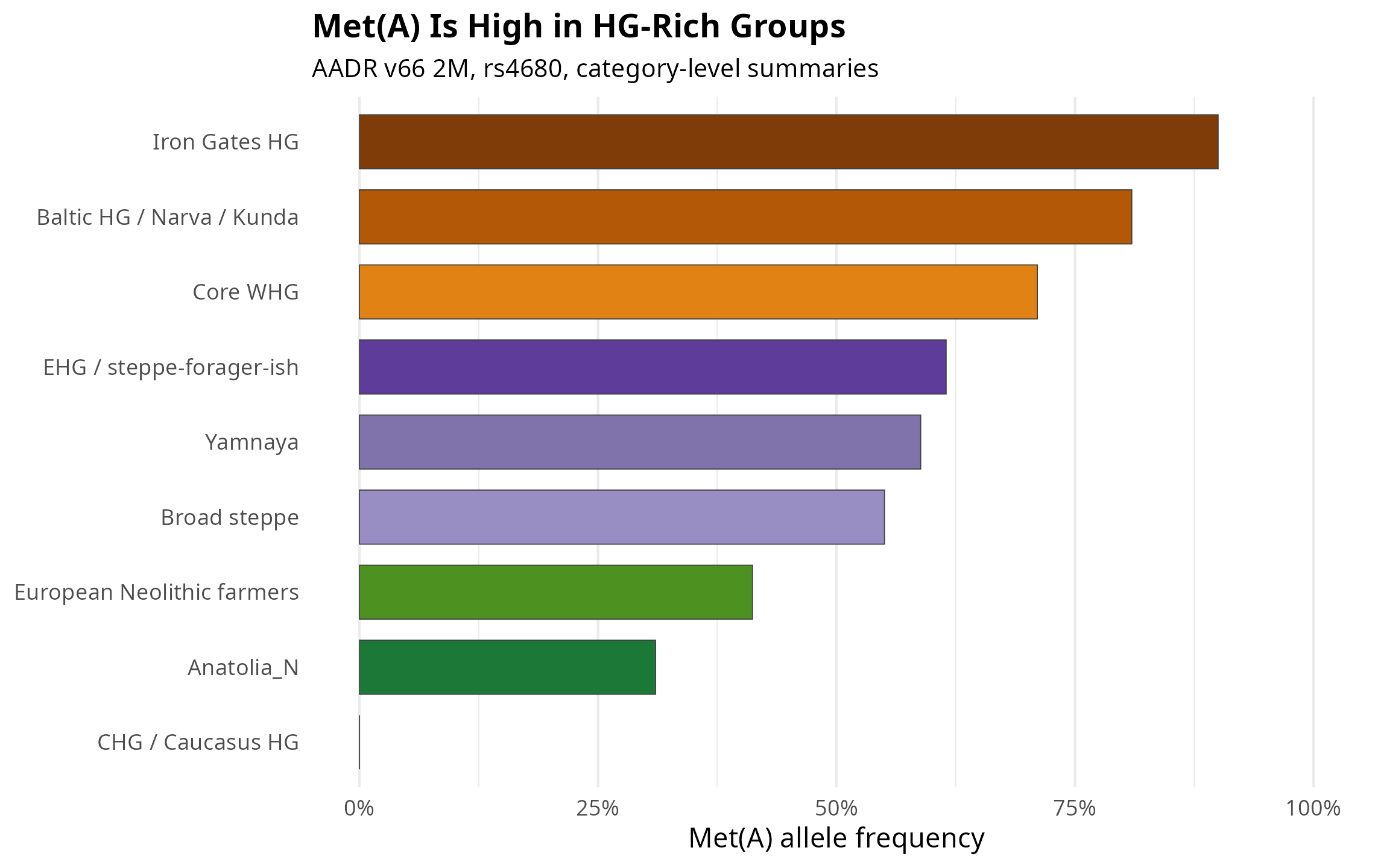
Table 2. Category-level allele frequencies. These labels are interpretive bins built from AADR Group_ID strings, so they should be read as descriptive rather than formal qpAdm classifications.
Upgrade to a paid subscription to unlock the full climate analysis, see the exact data models, and find out what really drove the evolution of the COMT Met(A) allele across deep history.